叶绿素怎么进化而来的?
这个问题也要分一下,因为至少有两种叶绿素:细菌叶绿素和高等植物里的叶绿素。
高等植物的叶绿素显然是细菌叶绿素进化而来的(包括基因突变等)
这一点我们很方便地找到进化上的证据。比如两者的叶绿素分子(负责光合作用的主要分子之一,结合光子)不太多,而且合成步骤几乎一样,除了 Protochlorophyllide 到 Chlorin 这一步。
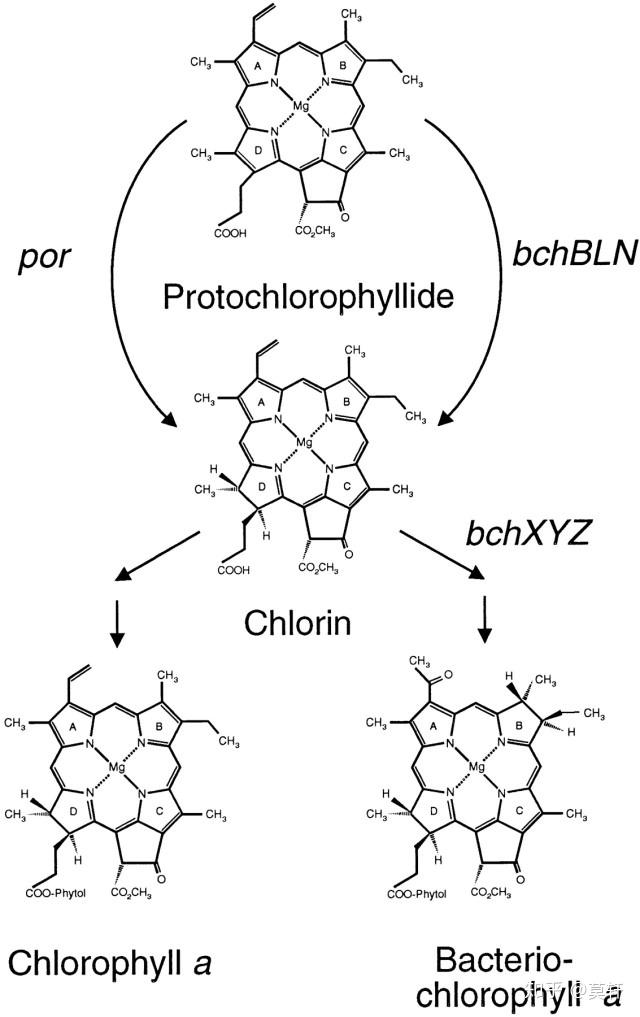
por 和 bchXYZ 的主要区别是,bchXYZ 对氧非常敏感,而 por 对光非常敏感。一个合理的假设就是,地球上的开始出现的富氧环境使得 bchXYZ,容易受到损伤,落伍了。同时,基因突变和光的定向筛选使得光敏感的 por 蛋白出现。(* 注:por 有三种,porA,porB,porC,对光的敏感度不一样),这点也是各位研究光合作用大佬的观点[1]。
那么,比叶绿素更早的细菌叶绿素怎么进化来的?我想这也是题主最想知道的。
顺着上文的思路,我们来稍微补一下地球化学历史。
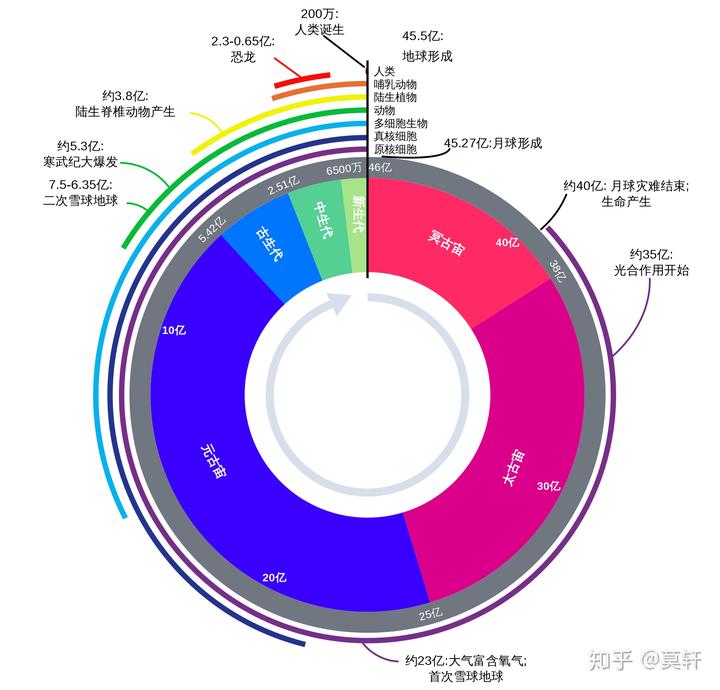
地球大约 46-50 亿岁,原始生命诞生约 40 亿年原始生命产生,约 35 亿年光合作用开始。
既然光合作用开始,那么负责吸收光子的叶绿素分子似乎也应该在这个时候登场。我们似乎应该去这个时期寻找答案,但问题是35 亿年前出了什么事情,光合作用怎么就一下子开始了?
普遍认为,这个时期原始生命将原始汤(前 40-35 亿年)中的丰富有机物消耗殆尽,生命开始出现第一轮危机,一些生命进化出利用光的能力,进行自养,而氧气作为副产品产生了。正是这个改变,导致了 10 亿年后一个重大事件的发生,大氧化事件。彼时,氧气在大气中的含量逐渐攀升至不可忽略的浓度。大氧化之后,依靠太阳能生活的细胞开始出现。
光合作用的产生自然离不开叶绿素,上文提到细菌的叶绿素分子是由 bchXYZ 合成。比较有意思的是,bchXYZ 竟然和现存的 Type I 型固氮酶 nifH, nifK, 和 nifD 氨基酸序列相似[2]。
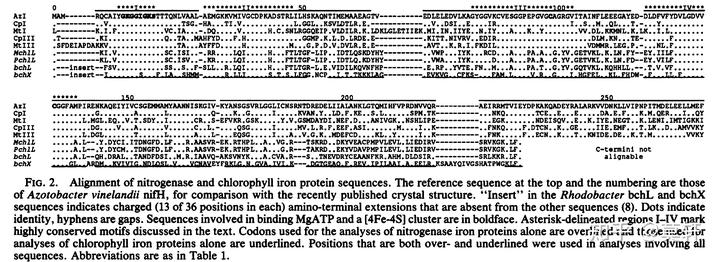
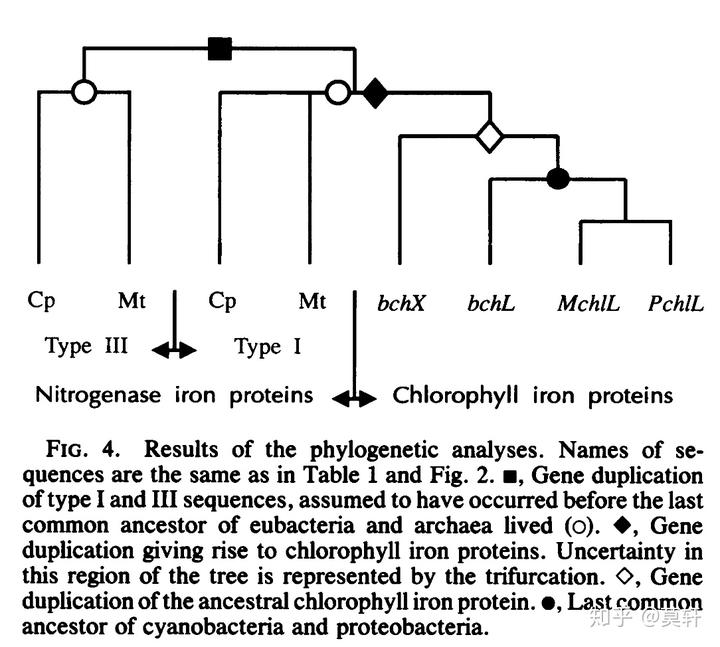
另外,bchXYZ 和 Type I 型固氮酶的另个一个相似性就是氧敏感性。总之,这些证据都表明,bchXYZ 是由固氮菌的固氮酶演化而来的。而始祖细胞第一批演化出来的就包含固氮菌和其他早期固氮生命[3]。
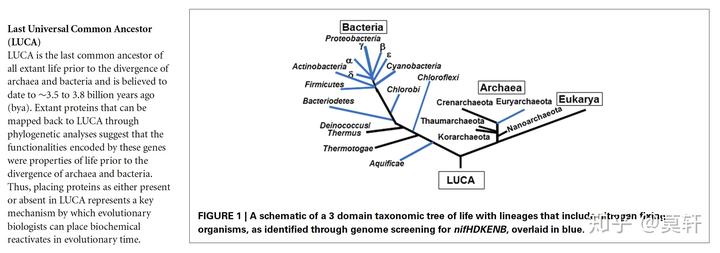
LUCA(Last Universal Common Ancestor)是普遍意义上认为的目前地球上所有生物的祖先,始祖细胞。
固氮酶将空气中的氮气转化成氨。固氮酶是一个非常有意思的酶,简单地说它有两部分构成,一个是蛋白部分,另一个是辅酶部分,单独任何一个部分都不表现出催化能力。以 nifH 为例,nifH 有一个铁钼硫基辅酶结构。nifH 和铁钼硫基辅酶结合以后,才能催化氮气分子形成氨。
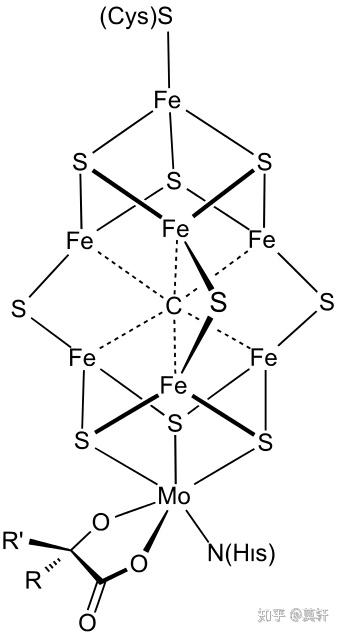
单独把 FeMoS 辅酶放出来,是因为,在地球早期的生命汤中,这个结构似乎是可以通过自然界的化学作用偶然地自发形成的。很有可能,在它形成之后结合不同的游离氨基酸(比如半胱氨酸或丝氨酸等)逐渐形成催化氮气分子的能力,进而加速了原始生命的诞生。虽然如此,在最初的十亿多年间,地球还是处在原始汤的状态。
这里假设性解释了负责细菌叶绿素分子合成酶的形成机制。
那么,总结一下就是:
原始汤中简单辅酶结构的形成—>逐渐形成简单的催化结构—>原始生命—>固氮菌——>光合作用细菌——>高等植物
相应地,作为酶的产物叶绿素就在这个复杂的过程中诞生。
第一个细胞(LUCA)如何能把产生的能量分子包住呢?
没有这些分子前不就是死物?
这两个问题,第一个通过化学键束缚。
譬如,叶绿素分子能够吸收一个光子,但这时候它仍然是叶绿素分子,只不过是能量比较高的叶绿素分子。在吸收光子之后,光子的能量被转化成了叶绿素分子中电子的势能。高能电子在释放之后,叶绿素就趋于稳定。同理,在 LUCA 中应该也有类似的能量约束机制,比如 Fe-S-Mo 基等。
当然,LUCA 的能量产生,肯定跟叶绿素和光合作用无关。
另外,高能能量分子存续时间并不久,都处在即产即消的平衡态。
没有这些分子前不就是死物?
什么是死,什么是活?
虽然原始汤中没有任何生命,但也旺盛地进行着随机或偶尔有序的化学反应,我觉得这也算生机。
一个原子核周围不断旋转的电子云,我们也可以认为是活的。因为,一个细胞放大以后,你看到的正是那些原子们(当然这有些诡辩),但正是这样诺大的集合才有了生命。
当然,你也可以认为 LUCA 之前都是死物,而 LUCA 之后才有了活物。
当然,看完我辛苦码完字的文章,动动手指点赞三连,大家都是活着的好胖友~
 微信扫一扫
微信扫一扫