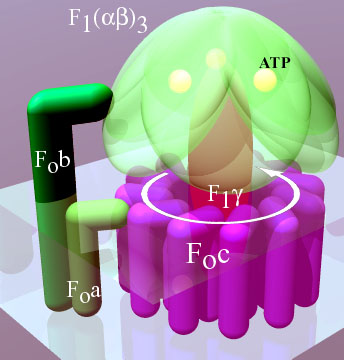
这是个常见的误解。生物体可以进化出轮子、轮轴那样的自由旋转构造,包括你在内的地球细胞生物身上经常遍布这样的构造(ATP 合酶),你的生存和身体运动极大依赖于 F 型 ATP 合酶通过转动进行的 ATP 合成。动物没有肉眼可见的“车轮”可归因于这种结构在自然地形上的效率低下。
 https://www.zhihu.com/video/1580197615710560256
https://www.zhihu.com/video/1580197615710560256
经常被人们提出的“轮子在演化上有解剖学障碍,轮子的静态部分和旋转部分之间的接口怎么处理”其实并不难办。扩散可及的小尺寸拼装结构的可动部分无需任何血管、神经连接,可动部分亦可由死细胞和细胞外基质组成。对于更大的结构,你可以参照现实存在的动物身上的自由旋转构造:
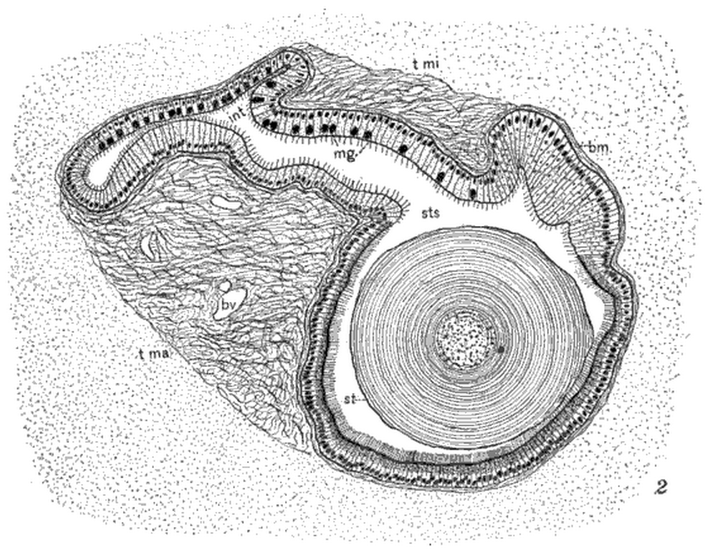
- 大部分双壳类和一部分腹足类的消化系统内有个结晶杆。它是在透明的糖蛋白外包裹着消化酶,由带有纤毛的囊不断分泌形成并延伸到消化道内,需要时可由纤毛推动让其突出端在消化道壁上旋转刮擦来磨碎食物并释放消化酶。不同动物的结晶杆的旋转速度有差异。在饥饿或干燥的环境里,一部分双壳类会消化掉结晶杆来应急。
这是肉眼可见的自由旋转构造,正如过去许多学者想象过的那样没有血管、神经之类不能跟着自由旋转的连接物。

细菌与古菌的鞭毛、真核生物的纤毛就是以旋转推动身体的细胞器官。地球上至少一半的细菌个体拥有 1 条或更多鞭毛,考虑到细菌压倒性的个体数量与物种数量,这意味着以旋转推动身体的移动方式其实是地球生物圈里最普遍、最常规的移动方式。
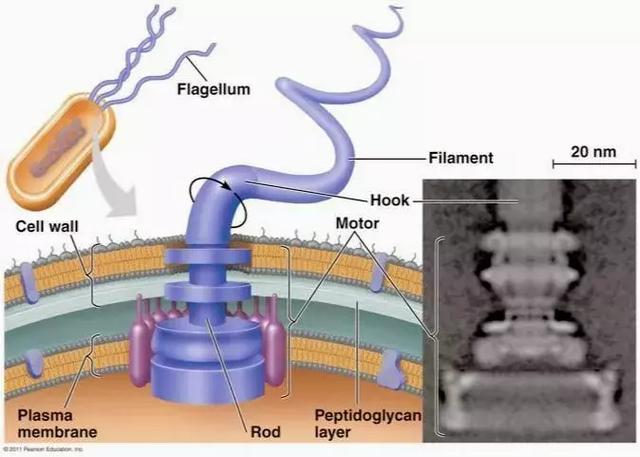
每个鞭毛由鞭毛丝、鞭毛钩、鞭毛马达组成。鞭毛丝是螺旋状的空心管,通过短而弯曲的鞭毛钩连接在鞭毛马达上,鞭毛马达是 20 个蛋白质分子组成的步进电机,可以顺时针或逆时针转动,其能源是电化学梯度下离子的跨膜流动,转动方向由磷酸化控制,可在毫秒内切换。细菌鞭毛的能量转换效率可以超过 60%;
纤毛是底部不旋转而尖端旋转的构造,能量转换效率同样很高。在多细胞生物中,栉水母至今还在以纤毛推进,该门级分类似乎已经这样延续了超过 6 亿年。一些栉水母的纤毛长达 2 毫米,如果你对螺旋桨的定义博爱一点,这就是地球生物身上的潜水艇螺旋桨了。
另外,古虫界副基体纲的一些单细胞真核生物的细胞上有一段区域可以连续旋转[1]。
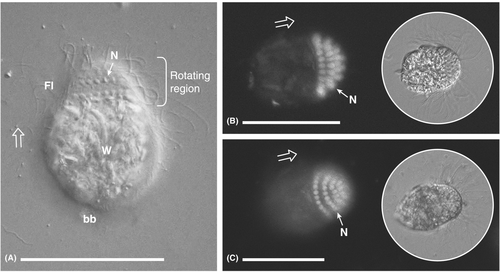
 https://www.zhihu.com/video/1696473349088387073
https://www.zhihu.com/video/1696473349088387073
对多细胞生物来说,“轮子与直升机螺旋桨”和多足步行·扑翼飞行相比毫无优势。旋转构造本身在发育上可以解决,但自然界缺少平整的铺装路面,直升机的能效比更是难看。不过,旋转移动跟轮轴、轴承并不绑定。地球上有许多全身卷成环状或抱成团来滚动的生物,人们早已观察到灰球菌属真菌、风滚草、轮蜘蛛、等足目、若干毛虫、虎甲幼虫、多足类、一些螳螂虾、潮虫、平头穴螈、南非犰狳蜥、穿山甲、刺猬、犰狳的滚动行为,乃至一些三叶虫化石表现出滚动的姿态,还有滚粪球的蜣螂。
- 以上效率问题可以被人自己佐证:南美洲土著知道轮子,他们能制造有小轮子的儿童玩具,但他们缺少大型家畜来拉动有意义的轮式车辆,所以他们的各种大规模工程建设都没有使用轮子。
如果地球上到处是极其平整的地面,你可以想象生物在蛋白质结晶或几丁质组成的轮轴与轴承的一侧长出一组负责分泌轮子或扇叶的腺体,分泌出蛋白质或几丁质的轮子 / 扇叶套在轴上,肌肉将其推出体外并驱动。滚珠和轮子 / 扇叶磨损后可以替换。
- 这种构造也可以为狂风呼啸的行星上自然诞生的生物组成风电机,让生物从气流中获取能量。
更简单的处理办法在罗伯特·索耶的《星丛》里展示过了:
- 你根本就没必要自己长个轮子,可以由共生生物组成你的轮子。
- 你可以参考轮椅的结构。
你可以设想,在一个地面极其平整的地球上诞生的人类会驯化一种擅长滚动的动物,将木厢架在数个这种动物之间做成四轮车代替马车、用这种动物拖动滑橇代替狗拉雪橇等。
 微信扫一扫
微信扫一扫