太长 / 太难不看版
因为运气不好,大概
(这篇回答主要针对“为什么没有已知生物呼吸氮气”,目前已经发现有很多不需要氧气的呼吸方式,所以答主认为问题的前半段与事实不符就没有解释)
地球大气中氮气占比的成因与生命活动关系不大
首先扯个皮,“远古地球大气中没有氮气,现今大气中的氮气是大氧化事件的产物”在现在是个与诸多观测结果相悖的假设,不适合作为回答本问题的论据
“远古地球大气中主要是氨等还原性气体”算是一个历史较久的设想,并且与著名的“原始有机汤”(Primordial soup)生命起源假说相伴相生。1953 年那个著名的的米勒 - 尤里实验的大气配方就或多或少受这一假设的启发:具体而言,斯坦利·米勒与哈罗德·尤里根据当时对木星等气体巨星的观测结果,推测地球早期的大气也将充满了氢、氨、甲烷和一氧化碳,这样还原性的气体
但越来越多的研究结果显示,远古大气的主要成分可能是氮气和二氧化碳等中性气体。采用铀 - 铅测年法[1]对热液石英中流体包裹体的成分分析表明,太古宙时期,氮气在大气中的分压及同位素比例已经与今天相差无几[2][3]所以“生命出现前的地球大气中没有氮气”的说法在目前并不是那么可信

氧气在呼吸作用中的角色是“电极”而非“助燃物”
回答与呼吸有关的问题,往往要涉及到两个母问题:什么是呼吸?氧气在其中扮演什么作用?
早在中学阶段,我们就接触过呼吸作用的三个阶段,以及发酵和无氧呼吸的概念。但中学教科书上很多便于记忆而简化的知识点给人一种错觉:最后那三十来个 ATP 的合成是氧气分子与辅酶[H]直接接触反应产生的,以及这些“呼吸”的过程并没有多大差别。然而事实上,现在的学术界对呼吸作用有一个更严格的定义:
将物质氧化释放的能量在电子传递链上制造膜内外电势差,并以这电势差驱动 ATP 合酶生成 ATP
若是依照这个定义,醋酸杆菌氧化乙醇产醋,酵母与植物产酒精,乳酸菌和人产生乳酸,这些中学课本上介绍的“无氧呼吸”都算不得呼吸作用。当然你可能会认为都是分解有机物产生 ATP,怎么就算不得呼吸作用,反倒要编那么一套叽里咕噜不讲人话?但学界那么分确实有它的道理:在上个世纪对细胞呼吸机理的艰难探索中,科学家们惊讶地发现,呼吸作用产生 ATP 的主要机制与以上这些生理过程,乃至与我们一般印象中的化学反应都大相径庭,反而更像是某些工厂里的机床

- 分解有机物释放的化学能或直接或间接(借助辅酶)地转入膜内侧的电子传递链上,并在此借由原电池反应转化为电势能驱动电子流
- 电能(电子流)又驱动电子传递链上的离子泵将氢离子往膜外侧泵出,制造膜内外离子浓度差
- 离子从高浓度侧流回低浓度侧时像河水滚水车那样以机械能转动 ATP 合酶,使之像磨盘一样把 ADP 与磷酸根“磨合”成 ATP
注意到了吗,驱动 ATP 合成的并不是哪个直接的化学反应,而是离子流回膜内侧时的机械能;而氧气在有氧呼吸中真正的作用,就是作为原电池的正极(最终电子受体),使电子传递链上产生巨大的电动势,继而增大单位时间内电子传递链上的电子流流量;而所谓的“无氧呼吸”,就是用氧气以外的其他物质作为最终电子受体的呼吸作用
而当明确氧气在呼吸作用中的位置后,我们的问题自然就变成了:取代这一位置的,可以是氮气吗?
目前并未发现有理化机制禁止氮气参与呼吸作用
首先指明,不是所有的呼吸作用的最终电子受体 / 供体都直接在电子传递链上反应。以我们熟悉的有氧呼吸为例,这一过程的负极或者说最终电子供体,是还原糖与脂肪等有机物,它们有时连膜都进不去,更遑论在电子传递链上直接反应;相对的,下文中提及的一些呼吸作用中,在电子传递链上直接反应的是则是最终电子供体
但不管怎样,呼吸作用产生能量的效率与电子传递链上的电动势直接相关,而后者又不会大于最终电子受体与供体之间还原能力的差值,因而我们希望有一种计量物质在特定条件下化学物质失去或获得电子能力的指标,这套指标被称为氧化还原电位(redox potential 或 oxidation / reduction potential,ORP),在组织细胞内,O₂/H₂O 的还原电位为 +0.815V。作为比较,我们来看看在其他呼吸作用中,各种最终电子受体的还原电位。你会发现在微生物的世界里,这电子受体的选择异常地多样[4]:
| 呼吸类型 | 最终电子受体 | 还原产物 | 还原电位(25℃, PH=7) |
|---|---|---|---|
| 高氯酸盐呼吸 | ClO₄⁻/ ClO₃⁻ | Cl⁻ | +0.797V |
| 铁还原 | Fe(III) | Fe(II) | +0.75V |
| 碘酸盐呼吸 | IO₃⁻ | I⁻ | +0.72V |
| 反硝化 | NO₃⁻ | N₂ | +0.40V |
| … | … | … | … |
| 富马酸盐呼吸 | 富马酸盐 | 琥珀酸盐 | +0.03V |
| 硫酸盐呼吸 | SO₄²⁻ | HS⁻ | -0.22V |
| 硫呼吸 | S | HS⁻ | -0.27V |
| 铀还原 | U(VI) | U(IV) | ?V |
| 产甲烷呼吸 | CO₂ | CH₄ | -0.25V |
| 乙酸化(产醋酸盐呼吸) | CO₂ | 醋酸盐(CH₃COO⁻) | -0.30V |
| … | … | … | … |
是的上面的表格直接扒的 Wiki 因为有氧呼吸一栏的氧气还原电位已经换算成细胞液环境下所以默认其他反应也已经换算过了,要是与实际数据有出入请大声地斥责我罢
在上表中,以二氧化碳为最终受体的两个反应对回答我们的问题具有重要的参考价值。在产甲烷呼吸中,二氧化碳的还原电位达到了 -0.25V,在产醋酸盐呼吸中则低至 -0.30V,甚至已经低于 N₂→NH₄⁺在中性水溶液中 -0.139V[5]的还原电位,这说明氮气在电化学上并不会是那么受生命嫌弃的选择
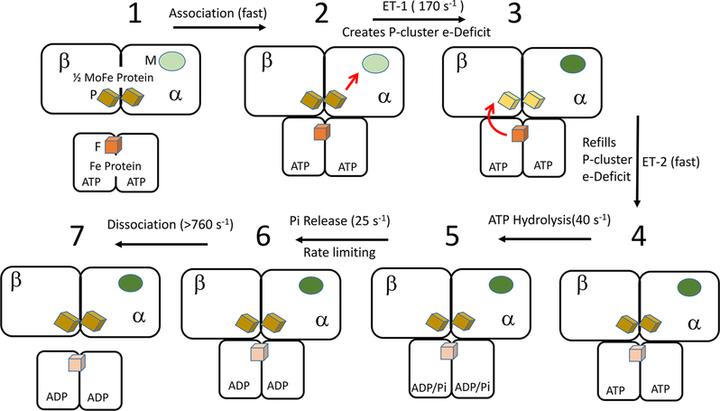
但会不会自发反应是一回事,反应难度又是另一回事,我们都知道固定氮的难点在于氮氮三键巨大的键能,但对比二氧化碳时我们注意到,破开 C=O 键所需的能量为 799kJ/mol,相比其他形式的分子键已经十分接近破开 N≡N 键所需的 942KJ/mol 能量[6];另外,现有的生命形式已经告诉我们,化学性质稳定并不是不可逾越的困难。在大约 28 亿年前,进化已经就氮匮乏的问题给出过一个答案:固氮酶(nitrogenase)是一类存在于部分细菌与古菌中的金属蛋白家族,能在常温常压下直接催化氨合成
为什么空气中 N 元素这么多,但没有一个动物进化到可以利用空气中的氮气合成氨基酸呢?
虽然现有的固氮酶存在着耗能高的问题:固氮酶固定一分子氮气需要 16 分子的 ATP,这意味着氧化一分子葡萄糖得到的那就有 28~38 个 ATP 中就有大约一半要折在里面。但近年来对固氮酶的研究揭示了一些有趣的东西[7]:固氮过程中的电子传递似乎早于 ATP 水解,换句话说,固氮作用中氮气分子是“自动”断键,固氮本身不需要 ATP 供能。那么在进一步的研究结果推翻它前,我们完全可以设想一个进化出不依赖 ATP,或者只需要少量 ATP 就能还原氮气的固氮酶的世界线
关于 ATP 在固氮作用中的角色,目前有理论认为它可能类似于一个“开关”:固氮作用涉及一个“铁蛋白循环”(Fe protein cycle),当与 ATP 结合时,还原态的铁蛋白被“开启”,像电磁铁一样吸附上钼铁蛋白为反应提供电子;而当与之结合的 ATP 被水解后,铁蛋白被“关闭”,继而解除吸附,直到下一次循环
综上所述,目前我们并未发现有任何机制禁止以氮气为最终电子受体的呼吸作用在进化中诞生,所以对题主的问题,咱也只能这么回答:
运气不好吧大概
If
虽然上文说并没有明确的机制禁止氮呼吸,氮气还原电位过低的问题使得即使氮呼吸存在,它也没有有氧呼吸效率来得那么高。但如果在哪个平行宇宙的地球上真的进化出了氮呼吸,那这地球上的生物可能实现了我们的固氮酶没做到的事——用尽量低的能耗去还原氮气分子
如果真是这样,那即使氮呼吸本身在未来的岁月中被边缘化乃至淘汰,这种低成本固定氮的机制却有可能被遗留下来:有了它,这条世界线上的生物能利用的氮资源要远远多于我们,这可能使得它们在进化中能诞生一些我们没有的特殊创新——比如炸药
进化出的爆炸物最初可能只是某种不太行的储能物质或者直接作为某种小分子活性物,在部分个体中大量淤积,但在捕食关系出现后可能就成为一种非常有效的防御手段,顺带装甲或骨骼也进化成容易产生破片的形式,成就一身谁也不想碰的一换一功夫;而如果这自爆能力再遇上真社会化、共生、性二型、变态发育、哺乳或者其他什么性状,就可能变成航母战斗群那样的巨大移动群落,几头有啥吃啥吸收倍儿棒的巨兽身上是层层叠叠的厚甲,厚甲的缝隙里是乳腺,大大小小的生物起起落落,咗几口乳汁就飞回 / 游回领地边缘,一见到入侵者就冲着眼睛鼻孔贴过去一通乱炸……一个枪林弹雨永不止息的生态系统,一个在文明之前就步入热兵器时代的世界


 微信扫一扫
微信扫一扫